

Эволюция слюнных желез позвоночных животных как результат

Плакоидная чешуя хрящевых рыб представляет собой пластинку с положенным на ней шипом. Пластинка лежит в кориуме вершина шипа через эпидермис выдается наружу. Вся чешуя состоит из дентина, образованного клетками кориума, вершина шипа покрыта эмалью, образуемой клетками базального слоя эпидермиса.
Зубная система гомодонтная. Зубы у ряда земноводных располагаются не только на альвеолярной дуге им, как и рыбам, свойствен полифиодонтизм.
Слюнные железы развиты лучше, среди них выделяют подъязычные, зубные и губные. Секрет желез уже содержит ферменты.
У ядовитых змей задняя пара зубных желез преобразована в ядовитые, секрет содержит токсины (яд).
Зубы гетеродонтные, т.е. дифференцированные: различают резцы (incisivi), клыки (canini), малые коренные (praemolares) и коренные (molares). У ластоногих и зубатых китов зубы не дифференцированы. Зубы сидят в альвеолах, на альвеолярных дугах челюстей основание зуба сужается, образуя корень.
Резцы и клыки весьма сходны с коническими зубами предков (рептилий), коренные зубы подверглись наибольшим эволюционным преобразованиям и впервые возникли у зверозубых ящеров.
Губы мясистые у сумчатых и плацентарных, подвижны, что связано со вскармливанием детенышей молоком. Губы, щеки и челюсти ограничивают пространство, называемое преддверием рта.
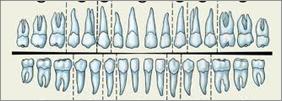
У человека зубная формула 2123
2123 (половина верхней и нижней челюсти).
Зубы, по сравнению с другими приматами, уменьшились в размерах, особенно клыки, они не выступают из зубного ряда и не перекрываются. Исчезли диастемы (промежутки между зубами) в верхних и нижних челюстях, зубы стали в плотный ряд, зубная дуга получила округлую (параболическую) форму.
Во время жевания нижняя челюсть может совершать вращательные движения по отношению к верхней, благодаря неперекрыванию редуцированных клыков и комплементарно расположенным буграм жевательных зубов обеих челюстей.
АТАВИСТИЧЕСКИЕ АНОМАЛИИ РОТОВОЙ ПОЛОСТИ ЧЕЛОВЕКА:
б) трехбугорчатые коренные зубы;
в) прорезывание сверхкомплектных зубов, т.е. у человек возможно образование большего, чем 32, количества зубных зачатков;
г) отсутствие «зубов мудрости»;
е) нарушение срастания (оно должно произойти к концу восьмой недели эмбриогенеза) костных горизонтальных складок которые образуют твердое небо, приводит к незаращению твердого неба и формированию порока, известного под названием «волчья пасть»;
Слюнные железы легко могут эволюционировать в ядовитые при правильных условиях
Ученые из Японии исследовали сеть генов, регулирующих выработку яда у змей и слюны у млекопитающих. Они показали, что эти гены высококонсервативны у самых разных видов и связаны в основном с модификацией, транспортом и деградацией белков. Для лучшего понимания эволюции ядовитости нужно исследовать не только гены токсинов, но и их регуляторную сеть.
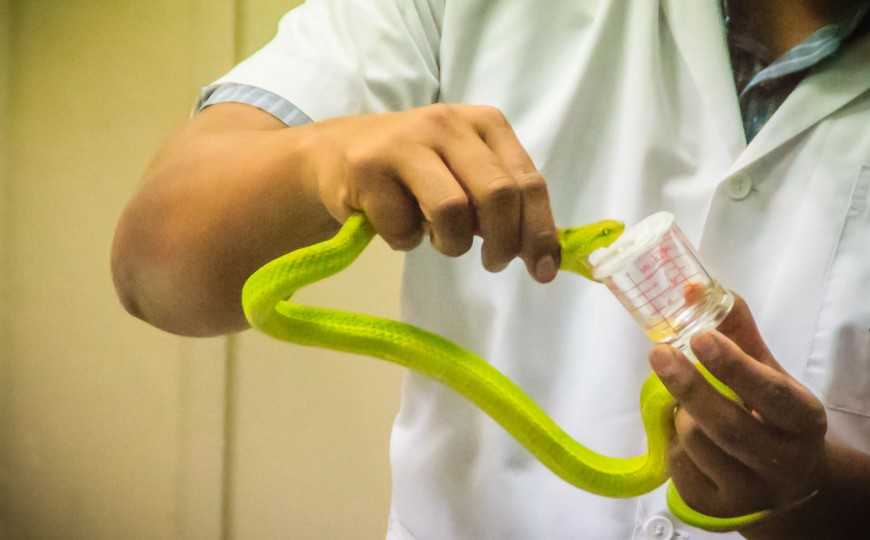
Способность к ядовитому укусу многократно возникала у позвоночных в ходе эволюции. Как и когда эта способность появлялась — остается загадкой. Большинство ученых начинают распутывать этот клубок с самого яда. Но его состав быстро изменяется, а у генов, вырабатывающих токсин, сложные паттерны экспрессии. Поэтому ученые из Японии подошли к проблеме с другой стороны, исследовав сеть генов, регулирующих выработку яда у змеи. Эту сеть они назвали metavenom network.
Ученые использовали РНК-библиотеки, составленные при изучении острочешуйной куфии (Protobothrops mucrosquamatus). Они анализировали гены, экспрессирующиеся совместно. Всего изучили 18 313 генов, которые разделили на 29 модулей размером от 38 до 3 380. Все гены токсинов попали в самый большой модуль 1, который и назвали metavenom network. Туда входят гены домашнего хозяйства, которые ассоциированы с выработкой яда. Паттерны их экспрессии уникальны для ядовитых желез. Гены, входящие в эту сеть, связаны в том числе с модификацией, транспортом и деградацией белков.
В модуль 1 входили также гены сигнальных путей UPR и ERAD, консервативных у многих животных. Ученые искали ортологи других генов, входящих модуль, у разных видов и нашли 546 ортологов у человека, шимпанзе, мыши, собаки, анолиса, жёлто-зелёной куфии, кобры, курицы и лягушки. Таким образом, гены из metavenom network высококонсервативны.
Далее авторы изучили транскриптомы слюнных желез мышей и неядовитых рептилий, таких как королевский питон (Python regius), маисовый полоз (Pantherophis guttatus) и пятнистый леопардовый эублефар (Eublepharis macularius). Кластеризация экспрессирующихся генов была очень схожа у этих транскриптомов и у первоначальной РНК-библиотеки острочешуйной куфии. На основе всех полученных данных ученые установили, что паттерны экспрессии сохранялись в гомологичных тканях, особенно если сравнивать ядовитые железы змей и слюнные железы млекопитающих.
Филогенетический анализ показал, что семейства генов из metavenom network эволюционировали быстрее у ядовитых змей, чем у других видов. В основном это были гены, связанные с модификацией и убиквитинированием белков, секрецией и перестройками хроматина.
Таким образом, ученые обнаружили сеть генов, которые регулируют как выработку яда, так и слюны. Они очень консервативны, но определить вовлеченность генов в конкретные процессы пока не удалось. Однако авторы предположили, что гены calr, manf, pdia6 и pdia3 работают как «контроль качества» нарабатываемых белков.
75. Эволюция ротовой полости у позвоночных.
Рассмотрим вначале эволюцию ротовой полости и ее производных. У бесчерепных ротовая полость окружена предротовой воронкой со щупальцами и частично выстлана мерцательным эпителием, который вместе с таким же эпителием глотки создает постоянный ток воды в кишечную трубку, несущую пищевые частицы и кислород. Ротовое отверстие позвоночных окружено кожными складками — губами, которые становятся подвижными только у сумчатых и плацентарных млекопитающих в связи со вскармливанием детенышей молоком.
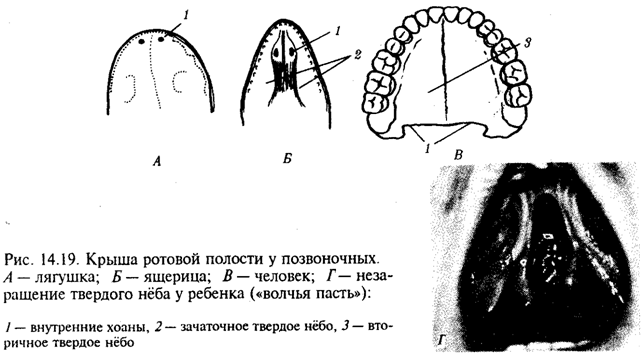
Крыша ротовой полости образована у рыб и земноводных основанием мозгового черепа, которое является первичным твердым нёбом. Хоаны земноводных открываются в их ротовую полость сразу позади альвеолярной дуги верхней челюсти. У пресмыкающихся объем ротовой полости увеличивается, и на верхнечелюстных и нёбных костях появляются горизонтальные складки, частично разделяющие ее на верхний, дыхательный, отдел и вторичную ротовую полость. Хоаны при этом несколько смещаются кзади. У млекопитающих наблюдается срастание этих складок по средней линии таким образом, что возникает сплошное вторичное твердое нёбо, полностью отделяющее друг от друга ротовую полость и полость носа. Хоаны при этом открываются в носоглотку. Этим достигается независимость функций органов ротовой полости от процесса дыхания (рис. 14.19, А— В).
До 7-й недели эмбрионального развития человека дифференцировка области stomodeum отсутствует. К концу 8-й недели происходит формирование вторичного твердого нёба за счет срастания горизонтальных костных складок. При нарушении адгезии клеток обеих складок возможно незаращение твердого нёба — порок развития, известный под названием волчья пасть (рис. 14.19, Г). Эта аномалия имеет атавистическую природу. Доказаны генетические механизмы ее возникновения. Она сопровождает ряд хромосомных синдромов, а также наследуется изолированно, причем с разной частотой у населения разных популяций. Так, в Японии частота «волчьей пасти» равна 2,1, а в Нигерии — 0,4 случая на 1000 рождений.
Зубы позвоночных связаны по происхождению с плакоидной чешуей хрящевых рыб (см. § 14.1). У них наблюдается непрерывный переход от типичных чешуи к зубам в ротовой полости (рис. 14.20). Зубы первоначально расположены во много рядов и покрывают всю слизистую оболочку ротовой полости, располагаясь у многих рыб даже на языке. У ряда земноводных зубы также расположены не только на альвеолярной дуге, но и на других костях, например на сошнике. У пресмыкающихся обнаруживается только один ряд зубов, причем, так же как у земноводных и рыб, дифференцировка их отсутствует. Такую зубную систему, в которой все зубы одинаковы, называют гомодонтной. У животных перечисленных классов зубы, как и плакоидные чешуи, могут многократно выпадать, сменяясь новыми их поколениями. Многократную смену зубов называют полифиодонтизмом.
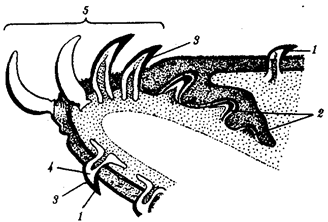
Рис. 14.20. Переход от плакоидной чешуи к зубам по краю ротового отверстия акулы: 1— плакоидная чешуя, 2—закладка новых зубов, 3—эмаль, 4—дентин, 5—зубы
Зубы млекопитающих дифференцированы на резцы, клыки и коренные. Они выполняют различные функции. Такую зубную систему называют гетеродонтной. Если резцы, и в особенности клыки, млекопитающих еще весьма сходны с коническими зубами предков, то наибольшим эволюционным преобразованиям подверглись коренные зубы. Впервые они вознили у зверозубых ящеров начала мезозойской эры за счет расширения оснований зубов и появления дополнительных бугорков, а затем и увеличения жевательных поверхностей при их сглаживании. Общее количество зубов у млекопитающих уменьшается и достигает у высших приматов 32. Зубы располагаются только на альвеолярных дугах челюстей, в ячейках. Основание зуба сужается, образуя корень.
Зубы человека по сравнению с другими приматами уменьшены в размерах, особенно клыки. Моляры имеют четырехбугорчатое строение. Зубная дуга округлой формы. В связи с дифференцировкой зубов увеличилась продолжительность их функционирования, в результате чего в онтогенезе сменяются только два их поколения: молочные и коренные. Это явление называют дифиодонтизмом.
У человека возможны атавистические аномалии зубной системы, связанные с нарушениями как дифференцировки зубов, так и с их количеством. Редкой аномалией является гомодонтная зубная система, в которой все зубы имеют коническую форму. Более частой патологией является трехбугорчатое строение коренных зубов. Нередко встречается прорезывание сверхкомплектных зубов в ряду или за его пределами, иногда даже на твердом нёбе (рис. 14. 21). Это свидетельствует о том, что у человека возможно образование большего количества зубных зачатков, чем 32, как это в норме встречается у низших млекопитающих и представителей более отдаленных классов позвоночных. Свидетельством тенденции к дальнейшему уменьшению количества зубов у человека является то, что нередко последние коренные зубы, так называемые «зубы мудрости», вообще не прорезываются, а если и прорезываются, то это происходит поздно — до 25 лет. Кроме того, эти зубы имеют явно рудиментарный характер, уменьшены в размерах и часто слабо дифференцированы.
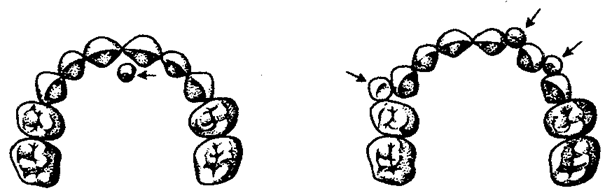
Рис. 14.21. Сверхкомплектные зубы, прорезавшиеся у человека (указаны стрелками)
На дне ротовой полости позвоночных располагается непарный выступ — язык, который у рыб представляет собой складку слизистой оболочки, лишенную мышц. Его движения осуществляются вместе с челюстями и подъязычной костью. У наземных позвоночных в языке располагаются мышцы, и они определяют его собственную подвижность. У пресмыкающихся и млекопитающих язык закладывается из трех зачатков: одного непарного и двух парных, лежащих кпереди от первого. Парные зачатки позже срастаются и дают начало телу языка. У большинства рептилий это срастание неполное, и язык раздвоен на конце. Весьма редким пороком развития языка у человека является раздвоенность его конца как результат несращения парных зачатков в эмбриогенезе.
Слюнные железы у рыб, заглатывающих пищу вместе с водой, отсутствуют и появляются только при переходе на сушу. Земноводные, обитающие на земле, имеют несколько слюнных желез, выделяющих только слизь. Слюна пресмыкающихся содержит уже и пищеварительные ферменты, а у некоторых змей—и токсины (см. §23.1). У млекопитающих слюнные железы многочисленны: это и мелкие — зубные, щечные, нёбные, язычные, гомологичные железам земноводных и пресмыкающихся, и крупные — подъязычные, подчелюстные и околоушные. Подъязычная и подчелюстная железы — результат дифференцировки подъязычной железы пресмыкающихся, околоушная — новое приобретение млекопитающих.
Эволюция начального отдела пищеварительной системы
Цель. Знать роль, эволюционные преобразования и основные этапы развития, а также строение органов начального отдела пищеварительной системы позвоночных. Знать и уметь обосновать формирование онтофилогенетически обусловленных пороков начального отдела пищеварительной системы человека.
Начальный отдел пищеварительной системы позвоночных связан с висцеральным черепом и включает в себя ротовую полость с расположенными в ней органами (язык, зубы, слюнные железы и др.), глотку, начальный отдел пищевода.
Эволюция ротовой полости позвоночных осуществлялась в направлении разделения первичной ротовой полости на пищеварительный и дыхательный отделы. У млекопитающих произошло разделение вторичной ротовой полости на преддверие и собственно ротовую полость.
Висцеральный скелет впервые появляется у первичных черепных, которые не имели челюстей, зубов и питались пассивно. Передний отдел их пищеварительной системы поддерживался околоротовым хрящевым кольцом с нерасчлененными хрящевыми висцеральными (жаберными) дугами, выполняющими функцию опоры. У современных челюстных животных жаберные дуги расчленились, что создало предпосылки для активного захвата пищи ртом и развития зубочелюстного аппарата. В дальнейшем эволюция висцерального отдела скелета у позвоночных животных проходила в направлении смены хрящевого скелета на костный, уменьшения количества костей, преобразования его элементов в структуры органа слуха (слуховые косточки) и опорного аппарата дыхательной системы.
Изменение среды обитания (выход позвоночных животных на сушу) привело к смене типа висцерального скелета, связанного с особенностями прикрепления челюстного аппарата к мозговому черепу: протостильный – гиостильный – аутостильный.
Появление и развитие зубочелюстного аппарата коррелировало с изменениями в висцеральном скелете и было связано с адаптациями к среде обитания и определенному виду питания: произошли укорочение длины челюстей, уменьшение количества зубов и их смены в течение жизни до двух (дифиодонтизм), дифференцировка зубов на резцы, клыки и коренные, изменялась форма жевательной поверхности коренных зубов и т.д. У человека, кроме того, в связи с изменением характера пищи и развитием речи зубная дуга приобрела параболическую форму, произошло смыкание зубов в единый ровный плотный ряд, уменьшилась длина клыков, исчезли диастемы, движение челюстей стало возможным в трех направлениях, лицевой угол приблизился к прямому, выдвинулся вперед подбородочный выступ.
Развитие слюнных желез позвоночных происходило от одноклеточных к многоклеточным, к их внеэпителиальной обособленности и расширению функций.
Рекапитуляция основных этапов филогенеза начального отдела пищеварительной системы наблюдается и в онтогенезе человека. Нарушение этого процесса приводит к развитию онто-филогенетически обусловленных пороков развития.
Задание для студентов
Работа 1. Функции начального отдела пищеварительной системы позвоночных
Изучите указанные функции и перепишите их в альбом.
1. Опорная – поддержание жаберного аппарата низших позвоночных.
2. Активный захват, удержание, перемешивание и измельчение пищи зубочелюстной системой.
3. Формирование пищевого комка; ферментативное переваривание пищи; защитная и регуляторная функции слюны.
4. Вкусовая рецепция пищи.
5. Участие в артикуляции.
Работа 2. Основные эволюционные преобразования начального отдела пищеварительной системы позвоночных животных
Используя материал лекций и методических пособий, изучите и перепишите в альбом основные эволюционные преобразования начального отдела пищеварительной системы позвоночных.
1. Смена функций: висцеральные дуги, выполнявшие функции опоры для жаберного аппарата, участвуют в образовании челюстей и слуховых косточек.
2. Усиление главной функции:
а) появление и развитие органов активного захвата пищи (челюсти, зубы, язык);
б) развитие единого зубочелюстного аппарата;
в) развитие слюнных желез по пути увеличения количества секретирующих клеток, их внеэпителиальной обособленности и дифференцировки; появление ферментативной активности слюны.
3. Разделение органов и функций:
а) разделение первичной ротовой полости на пищеварительный и дыхательный отделы;
б) разделение обшей гомодонтной зубной системы на зубы разного строения и функций.
4. Субституция: смена типа висцерального черепа в связи с изменениями условий обитания и питания (протостильный – гиостильный – аутостильный).
5. Расширение количества выполняемых функций: механическая и ферментативная обработка пищи, защитная, регуляторная, вкусовая рецепция, питание молоком матери, артикуляция, участие в образовании слуховых косточек среднего уха.
Работа 3. Строение начального отдела пищеварительной системы позвоночных
Используя материалы лекций, учебника, рис. 1 и методических указаний, изучите строение ротовой полости представителей различных классов позвоночных.
 Рис. 1. Строение начального отдела (ротовая полость) пищеварительной системы позвоночных:
Рис. 1. Строение начального отдела (ротовая полость) пищеварительной системы позвоночных:
а – миноги: 1 – кожные лепестки; 2 – ротовое отверстие. 3 – язык; 4;5 – роговые пластинки с зубами; 6 – внутренние губные зубы; б – костистой рыбы: 1 – зубы верхней челюсти; 2 – крыша ротовой полости (первичное небо); 3 – отверстие пищевода; 4 – жаберные щели; 5 – язык; 6 – зубы нижней челюсти; 7 – жаберные дуги; 8 – сошник; в – лягушки: 1 – зубы верхней челюсти; 2 – хоаны; 3 – глазные яблоки; 4 – отверстие слуховой трубы; 5 – отверстие гортани; 6 – нижняя челюсть; 7 – язык;
8 – отверстие пищевода; 9 – отверстие голосового мешка; г – рептилии: 1 – зубы верхней челюсти; 2 – сошник; 3 – хоаны; 4 – складки вторичного неба; 5 – отверстие евстахиевой трубы; 6 – отверстие пищевода; 7 – отверстие трахеи; 8 – язык;
9 – зубы нижней челюсти; д – человека: 1 – преддверие полости рта; 2 – твердое нёбо; 3 – мягкое нёбо; 4 – язык; 5 – резцы нижней челюсти; 6 – резцы верхней челюсти; 7 – клык верхней челюсти; 8 – малые жевательные зубы (премоляры) верхней челюсти; 9 – большие жевательные зубы (моляры) верхней челюсти; 10 – моляры нижней челюсти; 11 – премоляры нижней челюсти; 12 – клык нижней челюсти
Работа 4. Сравнительная характеристика висцерального черепа в ряду позвоночных
А. Используя материалы лекций, учебника, методических указаний, изучите и зарисуйте строение висцерального черепа позвоночных. Обозначьте основные структуры на рис. 2.
 Рис. 2. Строение висцерального черепа позвоночных:
Рис. 2. Строение висцерального черепа позвоночных:
Б. На основе материала учебника, лекций, рис. 1, 2, работ 3, 4 и приложения 1 заполните таблицу, используя представленные ниже характеристики ротовой полости и висцерального скелета.
Висцеральный скелет и его преобразования
Круглоротые Хрящевые рыбы Костные рыбы Амфибии Рептилии Млекопитающие
а) Ротовая полость первичная и хорошо выражена. Впервые появляются хрящевые челюсти с дифференцированной мощной жевательной мускулатурой.
б) Ротовая полость первичная и хорошо выражена. Челюсти образуются путем частичного окостенения хрящевой челюстной дуги и развития накладных и предчелюстных костей.
в) Ротовая полость первичная и хорошо выражена. Челюсти отсутствуют.
г) Ротовая полость частично разделена вторичным нёбом на вторичную ротовую полость и носоглоточный дыхательный отдел.
д) Вторичная ротовая полость делится на преддверие и собственно ротовую полость.
е) Обширная ротоглоточная полость, куда открываются отверстия хоан и евстахиевых труб, голосовая щель и вход в пищевод.
2. Висцеральный скелет.
а) Хрящевой, дифференцируется на предчелюстные (первую и вторую) дуги, челюстную, подъязычную и 5-7 пар жаберных дуг. Челюстная дуга выполняет функцию первичных челюстей, ее нёбно-квадратный хрящ – функцию верхней челюсти, а меккелев – нижней. Подъязычная дуга состоит из гиомандибулярного хряща (подвесок) и гиоида. Она фиксирует челюстную дугу к мозговому черепу и проводит звук к слуховой капсуле.
б) Костно-хрящевой. Верхняя челюсть состоит из покровных верхнечелюстных и предчелюстных костей. Она не связана с челюстной дугой. Нёбно-квадратный хрящ участвует в образовании костей первичного нёба. Нижняя челюсть образуется за счет частичного окостенения меккелева хряща и развития накладных костей – угловой и зубной. Подъязычная дуга частично окостеневает. Жаберные дуги образуют четыре пары окостеневших дуг, несущих жабры.
в) Костный. Верхняя челюсть образована сросшимися пред- и верхнечелюстными костями. Нижняя челюсть образована одной зубной костью, подвижно сочлененной с черепом. Верхний отдел челюстной дуги преобразуется в наковальню, а нижний – в молоточек, верхний отдел подъязычной дуги – в стремечко, а нижний – в передние рожки подъязычной кости. Первая жаберная дуга и ее копула преобразовались в тело подъязычной кости, задние рожки, а вторая и третья – в щитовидный хрящ.
г) Висцеральный скелет – хрящевой, представляет собой единый упругий каркас для жаберных мешков.
д) Костный. Сводчатое первичное нёбо. Вторичное нёбо образовано путем срастания нёбных отростков верхнечелюстных, межчелюстных костей и нёбных костей. Гиомандибулярный хрящ преобразовался в столбик, гиоид – в подъязычный аппарат, а жаберные дуги – в хрящи гортани.
е) Костный. Нёбно-квадратный хрящ окостеневает и вместе с накладными костями образует плоское первичное нёбо, которое является основанием черепа. Верхняя челюсть образована пред- и верхнечелюстными костями, нижняя челюсть – покровными костями и сочленовной костью нижнего отдела челюстной дуги. Гиомандибулярный хрящ превращается в слуховую косточку – столбик. Первая жаберная щель превращается в барабанную полость и евстахиеву трубу, а остальные зарастают.
Работа 5. Эволюция зубной системы позвоночных
Используя материалы лекций, учебника и методических указаний, заполните таблицу
 Работа 6. Эволюция слюнных желез позвоночных
Работа 6. Эволюция слюнных желез позвоночных
Используя материалы лекций, учебника и методических указаний, заполните таблицу
Характеристика слюнных желез и функции слюны
Эволюция пищеварительной системы в типе хордовые
Содержание:
| Предмет: | Зоология |
| Тип работы: | Реферат |
| Язык: | Русский |
| Дата добавления: | 01.07.2019 |
Если вам тяжело разобраться в данной теме напишите мне в whatsapp разберём вашу тему, согласуем сроки и я вам помогу!
По этой ссылке вы сможете найти рефераты по зоологии на любые темы и посмотреть как они написаны:
Посмотрите похожие темы возможно они вам могут быть полезны:
Введение:
Позвоночные, как и все животные, гетеротрофны и постоянно нуждаются в притоке органических веществ извне в виде пищи. Пища необходима для создания и обновления клеток и тканей, поддержания их химического состава и метаболических процессов. Кроме того, пища является источником энергии, расходуемой организмом на различные виды деятельности. Таким образом, питание является одним из важнейших компонентов общего обмена веществ и энергии организма с окружающей средой. Важная роль этой функции была в том, что общие принципы строения пищеварительной системы и физиологии питания сформировались очень рано в эволюции животных. В частности, структура и функции ферментных систем. Определяющие процессы пищеварения практически одинаковы у всех животных.
Морфологические принципы строения пищеварительного тракта, в которых выполняются различные стадии переваривания пищи, также распространены.
Фундаментальной особенностью всех представителей животного мира, грибов и большинства простейших является биотрофия, то есть обязательное использование в качестве источников энергии и материалов для построения клеточных структур органических молекул, синтезируемых другими организмами автотрофами (абиотрофами). Как правило, биологический материал, используемый в биотрофии, не может быть ассимилирован без предварительной деградации полимерных молекул до простых элементов, которые могут проникать во внутреннюю среду организма или клетки и не иметь видовой специфичности. Система процессов, обеспечивающих усвоение пищевых объектов, обозначается двумя не совсем совпадающими терминами: усвоение и питание и включает три основных процесса: усвоение пищи, переваривание и усвоение.
На ранних этапах эволюции позвоночных их пищеварительная система постепенно усложнялась, в ней появлялись новые органы. У всех современных животных от рыб до людей эта система построена по единому плану: за желудком следует тонкая кишка, в которой переваривается большинство видов пищи, также происходит всасывание, за тонкой кишкой следует большая кишечник, где процессы пищеварения и всасывания завершены. У позвоночных печень и поджелудочная железа имеют лучшие пищеварительные железы (моллюски имеют пищеварительные железы, часто пищеварительная железа одновременно выполняет функции печени и поджелудочной железы).
Пищеварительные железы являются наростами пищеварительного тракта; в процессе онтогенеза они превращаются в самостоятельные органы. Они поддерживают связь с тонкой кишкой с помощью протоков, которые открываются в кишечник. У позвоночных животных, в связи с их приспособляемостью к жизни в различных условиях окружающей среды и использованием разнообразных пищевых продуктов, развились их характерные особенности: сложная структура зубов, появляется многокамерный желудок (например, у жвачных животных), кишечный тракт (у травоядных) расширяется и т. д. тем не менее, у всех животных, от самых низших до самых организованных, химия пищеварения и участвующие в нем ферменты очень похожи. Таким образом, в ходе эволюции пищеварительная система постепенно усложнялась, добавлялись новые органы и, наконец, был разработан сложный механизм, который достигал наибольшей сложности у людей.
Эволюция пищеварительной системы хордовых. Природа питания и структура пищеварительной системы
У позвоночных эпителий пищеварительного тракта является энтодермальным на протяжении почти всей его длины. Очень слабая оральная вмятина приводит к формированию переднего эктодермального отдела (стомодеум), а совершенно незначительная вмятина на месте бывшего бластопора приводит к образованию заднего эктодермального отдела (проктодеум). Во взрослом государстве границы между этими отделами не могут быть установлены. Дифференциация кишечного тракта начинается с формирования переднего более широкого среза, из которого развиваются парные карманообразные выросты, растущие в направлении эктодермы кожного покрова, растущие вместе с его небольшими выпячиваниями и, наконец, вырывающиеся. Это жаберные мешочки, указывающие на некоторые передние, дыхательные, кишечные или глотки (глотки).
Часть кишечника, которая лежит перед этим отделом, является полостью рта (cavum oris). В задней части кишечника у наземных позвоночных брюшная стенка выступает в виде мочевого пузыря (амфибия мочевого пузыря), образуя у амниот большой аллантоис эмбрионального органа, который затем частично приводит к образованию мочевого пузыря взрослого животного. В этой задней части кишечника, которая называется клоака, выводные протоки эмбриональных почек также открыты.
У большинства позвоночных клоака сохраняется в зрелом возрасте; только у высшей рыбы мочеполовой тракт отделяется от задней части кишечника, а у живых млекопитающих выгребная яма эмбриона делится спереди назад на дорсальную часть прямой кишки и брюшной синус мочеполовой системы.
Часть кишечника, которая лежит между жаберной областью и выгребной ямой, уже является специальным пищеварительным органом. Различается на несколько отделов, различающихся как морфологически, так и по функции. Передняя суженная часть выстлана, как и отделы, которые лежат спереди, с многослойным эпителием и представляет пищевод (пищевод). Далее следует увеличенный разрез, часто изогнутый в форме буквы U, с однослойным железистым эпителием желудка (желудок). За самим желудком следует сам кишечник, также выстланный однослойным железистым эпителием; кишечник обычно образует более или менее значительные извилины и делится на «среднюю» или «тонкую» кишку позвоночных и «заднюю» или «толстую» кишку, на границе между которой иногда развивается более или менее значительный слепой выпячивание.
Эпителий желудка и самого кишечника является однослойным цилиндрическим. У личинки миноги, как и у Protopterus, весь кишечник выстлан двустворчатыми моллюсками, как у ланцета, с реснитчатым эпителием. У миног, рыб и земноводных в различных отделах кишечника наблюдаются остатки ресничного покрова.
Стенки кишечника состоят преимущественно из внутренней эпителиальной выстилки, покрытой слоем соединительной ткани с кровеносными сосудами. Это слизистая оболочка кишечника (слизистая оболочка). Слизистая оболочка покрыта более рыхлой подслизистой соединительной тканью (подслизистой), а затем мышечным слоем (мышечным), обычно состоящим из внутреннего слоя кольцевых волокон и наружного продольного слоя. Поперечно-полосатые мышцы обычно наблюдаются только в начальной части кишечника (полость рта, глотка и часть пищевода) и в конечной. Все остальные части пищеварительного тракта имеют гладкую мускулатуру.
Сокращения этой мышцы приводят к ритмичным движениям стенок кишечника или перистальтике, с помощью которых содержимое кишечника перемешивается и перемещается. Соединительная ткань и кишечные мышцы развиваются из висцерального листа (splanchnopleura) боковой пластинки. Из того же источника развивается наружная оболочка части пищеварительного тракта, лежащей в брюшной полости (часть пищевода, желудка и собственно кишечника), так называемая серозная оболочка, которая является не чем иным, как частью перитонеальной оболочки брюшной полости.
Поскольку эта слизистая оболочка образуется благодаря сходимости по средней линии висцеральных листьев парных боковых пластинок, естественно, что кишечник укреплен на двойных листьях брюшной брыжейки (mesenterium). Брыжейка брюшной полости по большей части исчезает, а дорсальная часть остается в виде брыжейки взрослого животного, подвешивая кишечник и подводя к нему кровеносные сосуды и нервы (ветвление блуждающего нерва и нервов симпатической системы).
В связи с пищеварительным трактом позвоночные имеют довольно значительную систему желез. В полости рта у наземных позвоночных развиваются более или менее сложные слюнные железы. В начальной части кишечника очень большая железа фактически развивается в виде полого непарного выпячивания, лежащего в брюшной брыжейке, печени. Не менее постоянной является другая большая железа, развивающаяся в одной и той же области кишечника из нескольких зачатков поджелудочной железы. Иногда у рыб в начале тонкой кишки все еще имеются железистые слепые выросты привратников, а в области задней кишки иногда наблюдается ректальная железа.
Кроме того, в качестве слепого выпячивания стенок кишечника также развивается гидростатический аппарат большинства высших плавательных пузырей рыб. У цистер и вдвойне дышащих рыб, а также у наземных позвоночных, парное выпячивание стенок задней части глотки вызывает воздушное дыхание органов.
Ротовая полость
Рот позвонка расположен на переднем конце тела, но он более или менее смещен в брюшную сторону. Особенно у селахий и осетровых, с развитием рыла, рот отодвигается вдоль брюшной стенки от переднего конца головы. Ротовая полость выстлана спереди эктодермой. Эпителий полости рта многослойный, частично кератинизирующий; у земноводных это ресничное. У амфибий в слизистой оболочке полости рта иногда имеется обильная сеть кровеносных сосудов; Поверхность слизистой оболочки увеличивается за счет образования складок, которые, таким образом, приобретают значение дополнительного органа дыхания.
Рот позвонка обычно ограничен кожными складками или губами, которые, однако, обычно неподвижны. Только у млекопитающих на губах развивается значительная мускулатура. Так они становятся мясистыми и подвижными. У форм, челюсти которых покрыты роговым покровом, образующим клюв, как у черепах, птиц и клоакальных млекопитающих, губы не развиваются. У млекопитающих заметная полость между губами и зубами развивает преддверие рта (расширяющееся по бокам многих обезьян и щечных мешочков).
У рта и амфибий крышка рта образована основанием самого черепа (первичное небо), которое по бокам дополнено небными и крыловидными костями. Внутренние ноздри или ханы расположены на крыше спереди.
Амниотическая полость рта разделяется развивающимся вторичным нёбом на верхнюю дыхательную секцию или носоглоточный проход (cavum naso-pharyngeum) и нижнюю секцию или вторичную полость рта (cavum oro-pharyngeum). Начало этого процесса наблюдается у ящериц, у которых горизонтальные складки появляются спереди и по бокам, простираясь от верхней челюсти. Эти складки твердого неба частично покрывают ханы. При спокойном положении языка внутри полости рта у ящериц и птиц его спина лежит как раз в промежутке между небными складками и полностью отделяет носоглоточный проход от самой полости рта. У крокодилов и млекопитающих нёбные складки сходятся вдоль средней линии и образуют полную перегородку вторичного неба.
У млекопитающих твердое небо продолжается в мягкое небо, которое имеет свою собственную мускулатуру и ограничивает полость рта от глотки до двойной складки слизистой оболочки, которая отстает. Мягкое небо распространяется вправо и влево в небные глоточные дуги (arcus palato-pharyngei), а в середине его у некоторых высших обезьян и у человека висит уступ.
На твердом небе млекопитающих обычно имеются плотные поперечные гребни, называемые небными пластинками. Эпителий небных пластинок часто имеет толстый роговой слой, который иногда значительно увеличивается (роговые пластины сирен и кит беззубых китов, который образует своего рода фильтрационную корку). По-видимому, небные пластинки помогают растирать пищу языком. У людей они также заложены и хорошо видны у новорожденных, но затем они уменьшаются.
В полости рта позвоночных развивается целый ряд органов специального назначения: зубы, язык и ротовые железы.
Зубы
У позвоночных края рта и часто часть внутренней стенки ротовой полости снабжены зубами, обычно конической формы, которые первоначально служат для удержания захваченной добычи во рту, а у высших позвоночных это часто используется для измельчения пищи.
Развитие зубов начинается с появления утолщения эктодермы, которое распространяется более или менее глубоко в соединительную ткань кожи. Это утолщение у селахий, рептилий и особенно у млекопитающих имеет вид непрерывного выроста, который называется зубной пластинкой. В основной части последнего происходит формирование зачатков отдельных зубов. Мезодермальные клетки формируют локальное утолщение сосочка, более или менее глубоко выступая в утолщение эктодермы. Клетки основного слоя эпителия расположены в виде колпачка, покрывающего сосок мезодермы. Этот колпачок состоит из цилиндрических ячеек, образующих на своей внутренней поверхности твердое вещество эмали и поэтому называется эмалевым органом. Наружные клетки сосков мезодермы расположены в правом слое, и они также образуют очень твердое вещество дентина на их внешней поверхности. Эти внешние клетки сосочка называются одонтобластами.
По мере увеличения массы дентина зуб, развивающийся таким образом, постепенно принимает свою окончательную форму и выталкивается, прорезая эпителий. В полностью сформированном зубе мы различаем очень твердый, богатый известью поверхностный слой эмали (субстанция адамантина), состоящий из призматических волокон, расположенных вертикально к поверхности зуба и покрытых тонким слоем органического вещества зубной кутикулой. Основная зубная масса состоит из дентина (субстанция eburnean), который имеет призматическую структуру с многочисленными ветвящимися канальцами, в которые отростки соседних клеток (одонтобласты) проникают в него в сосочках. Наконец, у млекопитающих основание зуба все еще покрыто снаружи слоем костного вещества, которое здесь называют цементом. Зубной сосочек хранится внутри зуба в виде зубной пульпы (пульпы), содержащей нервы и кровеносные сосуды, которые питают зуб.
Внутренняя часть дентина иногда содержит каналы с кровеносными сосудами (вазодентином) и очень похожа на кость, в которую он иногда проходит у основания. Эмаль иногда слабо развита (рыба и рептилии) или вообще отсутствует (некоторые рыбы и немного млекопитающих).
По мере износа зуба он ослабевает и выпадает, и его заменяют новыми, которые развиваются у низших позвоночных на протяжении всей жизни. Такая неограниченная смена зубов называется полифиодонтизмом. У млекопитающих количество изменений зубов ограничено, поэтому наблюдаются только две последовательные зубные системы (молочная и постоянная), это дипиодонтизм. Наконец, в некоторых случаях млекопитающие прорезают только одно, неизменное поколение зубов, которое называется монофиодонтизмом.
Форма зубов в простейшем случае является конической, и количество зубов может быть очень значительным. Селахи имеют очень разнообразные формы зубьев в виде гребней, трезубцев и т. д. в некоторых случаях они превращаются в плоские измельчающие пластины (особенно на склонах). Другие рыбы и низшие наземные позвоночные обычно имеют простые конические зубы. Иногда в этих зубах происходит радиальное складывание дентина, придающее зубу более или менее сложную внутреннюю структуру (чешуекрылых, много окаменелых чистящих ганоидов, стегоцефалов и некоторых рептилий). У млекопитающих происходит дифференциация зубов, увеличение их размеров и в то же время ограничение их количества. Задние зубы млекопитающих приобретают особенно сложную многотрубную или складчатую форму.
У млекопитающих зубы сидят только в челюстях, в отдельных клетках и сильно дифференцированы (система heterodont). Передние зубы резца имеют форму зубила и специально приспособлены для захвата и резки пищи, зубы, следующие за ними, сохраняют коническую форму, но часто они представляют собой значительные клыки, которые служат для разрыва пищи; задние зубы, лежащие по бокам челюстей, приобретают более сложную бугристую или сложенную форму и используются для измельчения пищи; это коренные зубы, передние зубы которых называются псевдокорневыми (praemolares), а задние настоящие коренные зубы; последние обычно более сложны, чем первые.
Зубы позвоночных, возникшие в виде чистых кожных защитных образований (чешуйчатые чешуйки), которые, кстати, также были обнаружены на краях рта, приобрели более значительные размеры на челюстях, где они играют роль вспомогательных органов для установки и удержания добычи.
Уже у примитивных позвоночных их форма часто изменяется в связи с изменением характера пищи; примерами являются режущие ланцетные зубы хищных акул и плоские шлифовальные зубы скатов и некоторых акул, которые питаются моллюсками. Обычно, однако, зубы имеют простую коническую форму и располагаются сначала на всех костях полости рта, а затем ограничиваются только одной челюстью. Такие зубы имеют неограниченное количество смен. У млекопитающих зубы подвергаются значительной дифференциации, поэтому развиваются как минимум три основных типа зубов, различающихся по форме и функциям: резцы, клыки и коренные зубы.
Зубы сидят в отдельных клетках, их основание сужается, образуя корень. Отдельные зубы увеличиваются в размерах, коренные зубы приобретают очень сложную форму, но количество зубов значительно уменьшается, и в то же время уменьшается число смещений, так что развивается не более двух поколений зубов, из которых только второе постоянное характеризует взрослое животное. С увеличением продолжительности функционирования таких зубов, особенно при значительном истирании, продолжительность роста увеличивается, так что иногда зубы растут на протяжении всей жизни.
Эта эволюция зубной системы, очевидно, обусловлена приобретением способности предварительно пережевывать пищу, что, в свою очередь, привело к ее быстрому и безупречному усвоению, что характеризует энергетический обмен и более интенсивную жизнедеятельность, как это наблюдается у млекопитающих. Можно сказать, что в эволюции последних, наряду с приобретением волос и бодрости, с развитием коры головного мозга, дифференциация зубной системы также играла важную роль.
Язык
В некоторых случаях это орган, построенный лишь немного выше, чем у рыбы; в большинстве случаев это гораздо более совершенный орган, по-видимому, имеющий более сложный состав; в любом случае, помимо складки, покрывающей переднюю часть подъязычного аппарата (рыбий язык), перед железом развивается специальное железистое поле. последний в нижней части ротовой полости, которая затем является частью языка. На языке наземных позвоночных, начиная с земноводных, появляется их собственная мускулатура, которая является продуктом выделения и дифференциации подъязычной мускулатуры.
Вкусовые органы языка иннервируются у наземных позвоночных глоссофарингеальным нервом.
У земноводных передняя часть языка изобилует железами, которые выделяют липкую слизь, которая помогает ловить насекомых и других жертв. В некоторых случаях язык может быть брошен далеко вперед. У бесхвостых амфибий язык обычно прикреплен к краю нижней челюсти, а свободный конец откидывается назад к горлу; во время рыбалки этот задний конец выбрасывается вперед. У земноводных, постоянно живущих в воде, язык сокращен.
Язык высших позвоночных, по-видимому, является морфологически более сложным образованием, основанным на данных об истории развития. Он развивается у амниот из неспаренного зачатка, лежащего в области второго и третьего жаберных арок, к которому затем прикрепляется еще пара валиков, лежащих по бокам, перед аркой подъязычной кости. Между этими зачатками лежит прокладка щитовидной железы. Задний зачаток дает начало корню языка, затем передние, парные зачатки сливаются друг с другом и дают начало телу языка.
В дополнение к двум ранее названным нервам, тройничный нерв также принимает участие в иннервации языка у рептилий. Форма и степень подвижности языка у рептилий сильно различаются. У черепах и крокодилов он имеет малую подвижность, а у большинства ящериц и змей он имеет удлиненную форму, обычно глубоко раздвоен в конце и чрезвычайно подвижен и чувствителен. В этом случае у основания языка имеется кольцевая складка, образующая влагалище языка. Сублингвальный аппарат образует скелет языка, а его передний непарный элемент (базихиал) распространяется на длинный отросток, поддерживающий переднюю часть языка (os entoglossum). Обе пары рогов подъязычного аппарата служат точкой прикрепления мышц, которые двигают язык. Хамелеоны имеют особенно длинный червеобразный язык, у которого на конце имеет железистое расширение.
У птиц обычно тонкий и малоподвижный язык лишен внутренних мышц, заострен спереди и характеризуется сильным развитием рогового слоя, который снабжен роговыми сосочками сзади языка. У дятлов и колибри очень длинный язык тянется далеко с помощью подъязычного аппарата, чьи задние рога огибают заднюю часть черепа и лежат на нем, достигая ноздрей. У попугаев широкий мясистый язык очень подвижен.
Язык млекопитающих развивается в целом аналогично языку рептилий. Собственная мускулатура достигает своего наибольшего развития, так что язык необычайно подвижен и выполняет множество функций, в частности он играет важную роль в акте глотания, а также как орган вкуса. Язык млекопитающих чрезвычайно богат железами и снабжен папиллами различных форм и значений, частично сенсорными (со вкусовыми рецепторами), частично ороговевшими. В целом, кератинизация языка иногда довольно значительна, как, например, у хищников. У муравьедов наблюдается очень длинный и липкий язык.
У некоторых сумчатых, полумах и обезьян есть язык (sublingua) под языком, который Гегенбаур сравнивает с языком низших форм, предполагая, что сам язык млекопитающих является новообразованием. Эта точка зрения не получила достаточного подтверждения.
Ротовые железы
У рыб и водных амфибий нет сложных ротовых желез. Последние представляют собой приобретение наземных позвоночных, у которых они первоначально служат для увлажнения слизистой оболочки полости рта и особенно для увлажнения пищи, но впоследствии у млекопитающих выделяемая слюна содержит не только слизь, но и серозную жидкость, которая включает, помимо прочего, пищеварительные ферменты, расщепляющие углеводы: птиалин, который превращает крахмал в мальтозу, и фермент мальтаза, который превращает мальтозу в глюкозу.

У птиц есть железы на небе и подъязычных железах, которые особенно развиты у грызущих птиц.
У млекопитающих устные железы достигают своего наибольшего развития. Помимо многочисленных мелких слизистых желез, таких как половые железы, щечные, небные и язычные, которые, по-видимому, сопоставимы с железами
То же название, что и у амфибий и рептилий, есть еще несколько крупных так называемых слюнных желез. Это:
Кроме того, у других млекопитающих обнаружены несколько других оральных желез, таких как молярные железы копытных и орбитальные железы собак.
В связи с полостью рта различные лимфоидные органы также наблюдаются у наземных позвоночных. Амфибии, рептилии и птицы имеют отдельные лимфатические узлы в разных местах стенки полости рта и особенно за хоаном. Млекопитающие имеют крупные лимфоидные образования в различных местах и в области миндалин (миндалины). Небные миндалины (tonillae palatinae) лежат у людей и в антропоидах между небно-глоточной и небно-язычной арками, по бокам глотки. У других млекопитающих они лежат в мягком небе.
Глотка
Раздел пищеварительного тракта, расположенный между полостью рта и пищеводом, называется глоткой. Его стенки выстланы многослойным эпителием и снабжены обильными поперечно-полосатыми мышцами. На его территории развивается весь жаберный аппарат, а также важные органы внутренней секреции щитовидной железы, эпителиальные органы и зоб.
Поэтому у рыб мы относим область жаберных щелей к глотке. У наземных позвоночных в глотке, а также иногда в полости рта соединены два пищеварительных и дыхательных тракта, которые разделены сзади. Помимо гортани, которая открывается на заднем конце глотки, перед ней также открываются отверстия евстахиевых труб, а у высших амниот, как правило, носоглоточные проходы.
Пищевод
Пищевод это часть кишечной трубки, которая уже используется исключительно для переноса пищи. Он не резко отграничен от следующего за ним отдела (только у млекопитающих установлена внешняя граница), но в целом отличается от него внутренней подкладкой с многослойным эпителием, таким как глотка и полость рта, и частично положением вне брюшной полости. полость и, следовательно, отсутствие серозной оболочки, что особенно характерно для высших позвоночных. В его стенках нет пищеварительных желез, а внутренний слой эпителия может даже ороговевать. Длина пищевода различна, и у наземных позвоночных она зависит от длины шеи, достигая очень большого значения у птиц. Внутренняя стенка обычно образует многочисленные продольные складки и часто обильно снабжена слизистыми железами. За слизистой оболочкой следует мышечный слой, который в передней части состоит из поперечно-полосатых волокон, в некоторых случаях образующих всю мускулатуру пищевода.
Желудок
Желудок обычно представляет собой увеличенный участок кишечника, который внешне довольно четко отличается от пищевода только у млекопитающих, но даже в этом последнем случае граница является в определенной степени условной, поскольку физиологическая граница между этими отделами может не совпадать с внешним видимым.
Физиологически желудок характеризуется как орган, в котором первая фаза пищеварения происходит в кислой среде. Его стенка выстлана слизистой оболочкой с сетчатым расположением складок. Он имеет однослойный цилиндрический эпителий и снабжен специальными трубчатыми пищеварительными железами. С этой точки зрения, некоторые низшие позвоночные, такие как ланцет, вообще не имеют желудка (циклостомы, двуглавые, двуногие и некоторые костистые рыбы, в частности карповые). Другие рыбы, такие как наземные позвоночные, имеют внешне более или менее четко различимый желудок. Иногда она вытянута или изогнута в форме буквы U, так что оба ее колена лежат параллельно. В целом, его форма тесно связана с общей формой тела животного: у животных с удлиненным телом оно вытянуто, лежит поперек и принимает форму сумки у животных с более коротким телом. Передняя стенка желудка в последнем случае вогнутая и образует так называемую малую кривизну (кривизна малая), а задняя выпуклая стенка образует большую кривизну (большая кривизна). Часть желудка, ближайшая к пищеводу, называется сердечной (лежащая в сердце), а часть, прилегающая к самой кишке, является пилорической. Последний отграничен от кишечника мышечной кольцевой складкой слизистой оболочки (пилорический клапан).
У рыб желудок обычно нечетко разграничен, хотя иногда он принимает форму довольно объемного блайнда за сумкой.
У амфибий и рептилий желудок отделяется гораздо четче, особенно у крокодилов, у которых также наблюдается дальнейшая дифференциация внутри самого желудка: за большим мешковидным железистым разрезом со значительной мускулатурой следует четко изолированная пилорическая часть.
У птиц такая дифференциация приводит к полной изоляции двух отделов переднего (сердечного) едва вздутого железистого желудка и мышечного желудка, разделенных четким сужением. Чрезвычайно толстые мышечные стенки последнего выглядят как два диска с сухожильными центрами, от которых вытягиваются радиальные волокна. Внутренняя стенка мышечного желудка снабжена железами, которые выделяют сплошную роговидную внутреннюю оболочку, которая иногда имеет бугристую поверхность. Эта твердая раковина вместе с проглоченной галькой служит для измельчения пищи (что крайне важно при отсутствии зубов у птиц). Мышечный желудок у гранвористых птиц достигает наибольшего развития. Иногда за мышечным желудком появляется небольшой привратник желудка.
Желудок млекопитающих характеризуется значительным развитием и дифференцировкой желез. Сердечные железы, дно и пилорика, несколько отличаются по своему гистологическому строению и характеру секрета. Из них последние выделяют слизистую, щелочную секрецию; нижние железы кислые. В нижних железах есть два типа железистых клеток: кубические, основные клетки, которые секретируют проэнзимы пепсиноген и прохимозин, и большие круглые покровные клетки, рассеянные среди основных клеток, которые секретируют соляную кислоту. Кроме того, нижние железы выделяют еще один фермент, липазу. Пилорические железы также выделяют пепсиноген и прочимозин.
Несмотря на довольно резкую внешнюю изоляцию желудка от пищевода, во внутренней структуре мы часто наблюдаем очень разные границы между отделами. Так, например, у клоакальных млекопитающих он выстлан тем же плоским слоистым эпителием, что и пищевод. У других млекопитающих та часть желудка, которая ближе всего к пищеводу, часто выстлана многослойным эпителием, и, таким образом, в желудке млекопитающего можно различить пищевод и собственно желудок, который, в свою очередь, распадается на области, характеризующиеся железами. специфичные для них сердечная часть, дно и пилорическая область.
Области желудка могут быть отделены друг от друга сужениями, и тогда желудок приобретает сложную форму. Иногда пищевод изолирован (в хомяке), в других случаях изолированы различные другие области желудка (дно и пилорическая область китообразных). Желудок наиболее труден у жвачных животных, в которых, как известно, он разделен на четыре части: объемный рубец с многочисленными сильными сосочками, сетка с клеточной внутренней поверхностью, книга с продольными складками в форме листьев и, наконец, удлиненные складки. сычуг с нижними продольными складками. Первые три секции выстланы многослойным эпителием, и только сычуг является настоящим желудком железистой природы (с дном и пилорическими железами). Желудок верблюда несколько проще, но также своеобразен: у рубца есть особые выпячивания в стенке водяных клеток, снабженные цилиндрическим эпителием и сердечными железами. В сети сычуг не изолирован. В водяных клетках верблюдов запасы воды сохраняются в течение длительного времени.
Кишечник
Весь кишечник на самом деле выстлан слизистой оболочкой с цилиндрическим однослойным эпителием, содержащим, помимо железистых бокаловидных клеток, также часто трубчатые пищеварительные железы. В этом отделе желудочно-кишечного тракта, как и в желудке, химическая обработка пищи (но в щелочной среде) происходит через пищеварительные ферменты, выделяемые железами, т.е. переваривание пищи, и, кроме того, поглощение веществ переводится в растворенный штат. Однако в некоторых случаях внутриклеточное пищеварение также наблюдается в кишечном эпителии позвоночных (у селахий и амфибий), и клетки выделяют амебоидные процессы и захватывают частицы пищи внутрь.
Важность поверхности кишечного эпителия в процессе абсорбции проясняет существование различных устройств, позволяющих удлинить путь, пройденный едой, и увеличить поверхность всасывания путем образования складок и ворсинок.
У низших позвоночных это достигается формированием одной большой складки, спиральной вдоль большой части кишечника и называемой спиральным клапаном. У миног такая складка низкая и тянется вдоль всего кишечника, образуя едва заметный спиральный изгиб. У Селахии складка глубоко врезается в просвет кишечника и образует более или менее крутые повороты, напоминающие спиральную лестницу. Внутренние края клапана обычно изгибаются так, что его обороты более или менее перекрывают друг друга конусами. В некоторых случаях обороты клапана очень наклонные, а его свободный край плотно скручен сам по себе. У ганоидов и у рыб с двойным дыханием клапан обычно менее развит, а у костных ганоидов (чешуекрылых) его можно назвать рудиментарным. У костистых рыб нет спирального клапана (исключение Chirocentrus). При наличии спирального клапана кишка имеет относительно небольшую длину, но большой диаметр; еда проходит по такому кишечнику по спирали вдоль клапана.
Вместо этого у высших рыб, а также у наземных позвоночных наблюдается расширение самого кишечника, которое в то же время образует более или менее значимые извилины. У разных позвоночных длина кишечника очень различна, и это часто находится в явной связи с природой пищи; Как правило, можно признать, что у травоядных форм эта длина более значительна.
Увеличение поверхности всасывания кишечника достигается, кроме того, путем образования складок и ворсин. У низших рыб обычно наблюдаются продольные складки слизистой оболочки, обеспечивающие, в основном, возможность расширения кишечника. У высших рыб, а также у земноводных и у рептилий развивается нерегулярная сеть из многочисленных перемычек, которые иногда придают клеточную структуру внутренней оболочке кишечника. Края ригелей часто оказываются неровными, зазубренными и, если выемки глубоко прорезаны. Тогда таким образом развиваются неправильные ряды многочисленных, более или менее высоких сосочков. Это можно рассматривать как первые признаки появления кишечных ворсинок, массы которых покрывают кишечник птиц и млекопитающих и придают ему бархатистый вид.
Кишечник позвоночных обычно более или менее четко разделен на отделы. Его передняя часть, в которую протекают протоки печени и поджелудочной железы, изолирована у высших позвоночных под названием двенадцатиперстная кишка (duodenum), в которой имеются железы (Brunner’s), вблизи пилорических желез желудка. Обычно у низших позвоночных животных, помимо основной части, они различают «среднюю» кишку, которая содержит многочисленные либеркуновые железы у наземных позвоночных, заднюю или толстую кишку, которая у некоторых ганоидов и костистых рыб, особенно у Наземные позвоночные, ограничены кольцевой складкой (valvula ileo-colica). Высшие позвоночные на границе между тонкой и толстой кишкой обычно имеют более или менее значительный слепой вырост (коэкум). Толстая кишка, как правило, довольно короткая, у млекопитающих достигает более значительной длины и, в свою очередь, может быть разделена на саму кишку (ободочную кишку) и прямую кишку. Внутренняя поверхность толстой кишки обычно гладкая, а ее задняя часть называется клоакой, если в ней открыты протоки почек и половых желез. Брюшная стенка выгребных ям у земноводных образует саккулярный отросток мочевого пузыря, который у эмбрионов высших наземных позвоночных имеет большое значение как аллантоис, который служит не только резервуаром для мочи, но также и эмбриональным органом дыхания. У рептилий, птиц и млекопитающих он сильно растет, но к концу эмбриональной жизни большая часть его вырождается, и лишь очень небольшая его часть дает мочевой пузырь взрослого животного.
По всему кишечнику у разных позвоночных наблюдаются различные наросты желез. Протоки двух крупных пищеварительных желез печени и поджелудочной железы поступают в переднюю часть среднего кишечника у всех позвоночных. В той же области более высокая рыба часто имеет большее или меньшее количество (от одной до двухсот или более) слепых выростов, называемых аппендиксами pylorici и служащих, по-видимому, только для увеличения поверхности всасывания. Селахи в передней части задней кишки имеют железистый придаток в форме пальца, ректальную железу, функция которой неизвестна.
В результате активности всех этих желез кишечный сок содержит щелочную слизь со многими пищеварительными ферментами (у млекопитающих они различают эрепсин, затем липазу, амилазу, инвертазу, мальтазу, лактазу и, кроме того, энтерокиназу, которая активирует трипсиноген поджелудочной железы, превращая его в трипсин).
Пищеварительные железы
Печень
Печень это огромная железа, которая сочетает в себе несколько совершенно разных функций. Это пищеварительная железа, выделяющая желчь в кишечник, которая благодаря своей щелочной реакции нейтрализует желудочный сок и, кроме того, эмульгирует жиры, активирует липазу панкреатического сока и, следовательно, способствует расщеплению жиров, растворяет жирные кислоты и стимулирует кишечная моторика. В то же время печень является чрезвычайно важным очищающим кровь органом, который нейтрализует вредные продукты распада и образует, среди прочего, мочевину и мочевую кислоту, и, наконец, она служит резервным резервуаром для углеводов, которые откладываются в ней. как гликоген.
Печень развивается из направленного вперед слепого выроста брюшной стенки кишечника, лежащего в брюшной брыжейке. Ветви зародыша печени имеют древовидную форму, приобретая все более сложную структуру. У cyclostomes и у selachia эти ветви сразу полые; у других позвоночных они сначала развиваются в виде компактных клеточных нитей, которые только позже могут канализироваться. Между ветвями мезенхима проникает в кровеносные сосуды, и общая масса железы быстро увеличивается. По способу его развития печень представляет собой сложную трубчатую железу. Однако в будущем между ветвями развиваются многочисленные анастомозы, а печень приобретает характер ретинальной железы.
Печень взрослого животного лежит позади сердца в желудке. Это большой двухлопастный или многолепестковый орган, содержащий сложную сеть желчных капилляров, расположенных между клетками печени и соединенных в более крупные желчные протоки, в конечном итоге протекающих в выводные печеночные протоки (ductus hepatici). Печеночные протоки обычно соединены в один канал, снабженный слепым выростом желчного пузыря (vesica fellea), который служит резервуаром, в котором накапливается желчь. Узкая шейка мочевого пузыря называется пузырным протоком (ductus cysticus), а общий проток от мочевого пузыря до слияния начальной части средней кишки обозначается как желчный проток (ductus choledochus). Иногда, кроме того, существуют также печеночные протоки (d. Hepatoenterici), протекающие в кишечник независимо от мочевого пузыря и общего протока. В некоторых случаях желчный пузырь отсутствует.
Печень получает для своего питания артериальную кровь через печеночную артерию, но, кроме того, получает большое количество венозной крови через портальную систему печени (v. Portarum hepatis).
Форма печени соответствует общей форме тела животного и, кроме того, зависит от положения и формы соседних органов. У млекопитающих он прилегает к диафрагме спереди. Обычно он делится на две правые и левые лопасти, которые, в свою очередь, могут распадаться на лепестки разных размеров.
Общий размер печени у низших позвоночных (рыбы и амфибии) относительно больше, чем у высших. У плотоядных животных печень крупнее. Чем травоядные.
Так как печень развивается в брюшной брыжейке, у взрослого животного она приостановлена на остатках этой брыжейки, известных как связки, на брюшной стенке тела (lig. Suspensorium печеночная) и на кишечнике (lig. Hepato-entericum ), а у млекопитающих к диафрагме.
Как показывает история развития, печень возникла из простого железистого выпячивания брюшной стенки кишечника, который, постепенно усложняясь, превратился сначала в разветвленную трубчатую железу, а затем в ретикулярную. Сравнение подтверждает этот вывод. У ланцета нет печени в собственном смысле этого слова, но есть железистый слепой вырост, направленный к брюшной стенке кишечника, в стенке которого сильно разветвлен венозный сосуд, несущий кровь из кишечника и, очевидно, соответствует воротной вене печени. Несомненно, этот нарост представляет собой рудиментарную печень позвоночных. У миксинов печень все еще сохраняет первичную природу разветвленной трубчатой железы (у миног желчные протоки и протоки разрастаются, а печень сохраняется только как эндокринная железа). У всех позвоночных он имеет характер сетчатки.
Поджелудочная железа
Поджелудочная железа является очень важной пищеварительной железой. Выделенный им сок содержит различные ферменты, которые расщепляют белки и углеводы. Кроме того, это, однако, и эндокринная железа.
Поджелудочная железа развивается из ряда мелких почек, которые затем образуют несколько зачатков в начале среднего кишечника в той же области, где лежит печень. Чаще всего имеется один спинной отросток и два брюшных, которые затем соединяются вместе в брыжейку позвоночника.
У взрослого животного поджелудочная железа более компактна. Это орган, состоящий из рассеянных долек, некоторые из выводных протоков которых (ductus pancreatici) втекают в начальный участок среднего кишечника, иногда независимо друг от друга, иногда предварительно соединяясь в один проток или впадая в общий желчный проток.
Поджелудочная железа имеет структуру сложной альвеолярной железы и обычно располагается в петле начального отдела среднего кишечника (двенадцатиперстной кишки птиц и млекопитающих). В некоторых случаях он спрятан в стенках кишечника (у миног и у двуногих животных), разбросан в виде небольших долек в брыжейке позвоночника (у некоторых костистых рыб) или встроен в ткани печени (у миксины и у некоторых). костистые рыбы). У млекопитающих это довольно компактный орган, лежащий в петле двенадцатиперстной кишки, в которую он обычно впадает в два протока брюшного, основного (ductus Wirsungi) и дорсального, дополнительного (d. Santorini). У артиодактилей только последнее иногда сохраняется.
Сок поджелудочной железы содержит ряд пищеварительных ферментов: трипсиноген (который дает трипсин с энтерокиназой), эрептазу, диастазу и липазу. Между ветвями самой поджелудочной железы расположены группы клеток, снабженные широкими кровеносными капиллярами островков Лангерганса. Эти островки играют роль желез внутренней секреции, имеющих большое значение в углеводном обмене.
Заключение
При всем разнообразии строения и биологии отдельных классов на примере амниот можно увидеть основные направления прогрессивной эволюции хордовых. Эти направления в наиболее общем виде характеризуются развитием структур и функций, повышающих общий уровень обмена веществ и биологической активности. В самой прямой форме это связано с эволюцией газообмена, а также системы кровообращения и пищеварения. Не менее важным является направление эволюции, связанное с разработкой систем регуляции внутриорганизма и координации функций, в конечном итоге направленных на повышение автономности организма, его способности нормально функционировать в условиях сложной и динамичной внешней среды. Это направление характеризуется эволюционными изменениями основных регуляторных систем организма центральной нервной и эндокринной систем.
Питание является основой энергетического и пластического обмена. Поэтому активация питания, повышение эффективности усвоения пищи представляют собой основную линию прогрессивной эволюции всех животных, включая позвоночных. Решающее значение функции питания определялось тем фактом, что в эволюции животных биохимические принципы процесса пищеварения развивались очень рано; Совокупность пищеварительных ферментов, их химический состав, специфичность действия характеризуются большим сходством в различных таксонах животного мира.
В ходе эволюции позвоночных морфологические принципы повышения эффективности пищеварения основывались, во-первых, на морфофункциональной дифференцировке первичной пищеварительной трубки и, во-вторых, на увеличении поверхности контакта эпителия кишечника с пищевой массой, которая В частности, интенсифицируется процесс усвоения и, соответственно, степень усвоения содержащихся в пище питательных веществ.
Позвоночные животные характеризуются собственными структурными особенностями и функциями пищеварительной системы. У большинства беспозвоночных сам процесс пищеварения, т.е. ферментативное расщепление сложных молекул пищевых веществ на простые компоненты, которые могут проникать через стенку кишечника и попадать в кровоток, осуществляется только в определенном отделе пищеварительного тракта. Здесь весь комплекс ферментов действует непосредственно на пищу, и здесь в большинстве случаев также происходит всасывание. Остальная часть кишечной трубки выполняет чисто транспортную функцию.
Позвоночных животных отличает сочетание морфологической и функциональной дифференцировки пищеварительного тракта. Пищеварительная система этих животных представляет собой своего рода многофункциональный «конвейер», во время которого в разных морфологически различных отделах проводятся последовательные стадии переваривания пищи от глотания пищи до поглощения питательных веществ и удаления непереваренных остатков.
Присылайте задания в любое время дня и ночи в  whatsapp.
whatsapp.
Официальный сайт Брильёновой Натальи Валерьевны преподавателя кафедры информатики и электроники Екатеринбургского государственного института.
Все авторские права на размещённые материалы сохранены за правообладателями этих материалов. Любое коммерческое и/или иное использование кроме предварительного ознакомления материалов сайта natalibrilenova.ru запрещено. Публикация и распространение размещённых материалов не преследует за собой коммерческой и/или любой другой выгоды.







