Нервные импульсы
Автор работы: Пользователь скрыл имя, 07 Апреля 2014 в 01:30, доклад
Краткое описание
Вся информация в мозг поступает от органов чувств — глаз, ушей, носа, языка и кожи, — а также от рецепторов растяжения в мышцах и терморецепторов, которыми усеяно все тело. Их можно сравнить с устройствами ввода компьютера. Данные поступают в особые входные, или сенсорные, области полушарий мозга.
Сигналы, передаваемые нервами, называют импульсами. Они очень похожи на электрические сигналы в схеме компьютера. Каждый нейрон обладает крошечным электрическим зарядом даже в состоянии покоя.
Вложенные файлы: 1 файл
Движение нервного импульса.docx
Категория: Человек, Автор: VseZnamus
Вся информация в мозг поступает от органов чувств — глаз, ушей, носа, языка и кожи, — а также от рецепторов растяжения в мышцах и терморецепторов, которыми усеяно все тело. Их можно сравнить с устройствами ввода компьютера. Данные поступают в особые входные, или сенсорные, области полушарий мозга.
Сигналы, передаваемые нервами, называют импульсами. Они очень похожи на электрические сигналы в схеме компьютера. Каждый нейрон обладает крошечным электрическим зарядом даже в состоянии покоя.
Импульс, поступивший, скажем, от глаза, вызывает движение определённых химических веществ к нейрону, в результате чего по аксону проходит волна электрического заряда — подобно тому, как по проводу движется электрический ток. На кончике аксона происходит выделение этих веществ, приводя к возбуждению близлежащих нейронов. Таким способом поступивший от глаза сигнал попадает в зрительный центр головного мозга. Здесь он обрабатывается, после чего посылаются ответные сигналы.
Импульсы, передаваемые нейронами всех типов, в основном сходны между собой. Каждая часть мозга расшифровывает входящие сообщения исходя из того, сколько их поступает за один раз. Например, сигналы от руки, касающейся чего-то горячего, будут поступать значительно интенсивнее сигналов от уха, которое в ту же минуту слышит пение птиц. Таким образом мозг определяет, какая информация для организма важнее.
Нервные импульсы проходят через мозг со скоростью до 400 км/ч, и каждую секунду через нейрон может пробегать 200 сигналов. В отличие от компьютера, который можно выключить, этот «управляющий» человеческого организма работает круглые сутки. Даже во время сна в мозг и из него ежесекундно поступает 50 миллионов нервных сигналов, и расходуемой при этом мощности хватило бы на питание 10-ваттной лампочки.
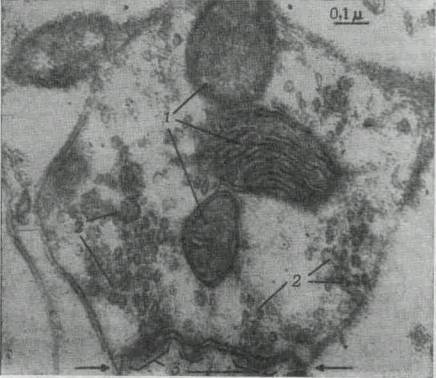
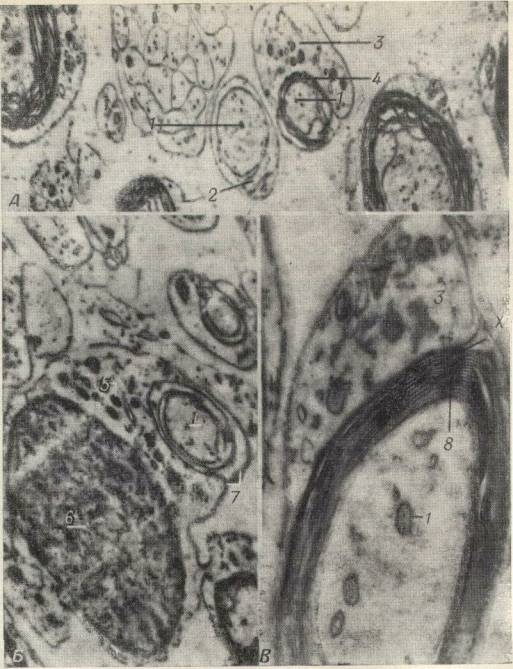
Нервные центры состоят из множества нейронов, связанных между собой еще большим множеством синаптиче ских связей. Это обилие синапсов определяют основные, свойства нервных центров: односторонность проведения возбуждения, замедление проведения возбуждения, сум-мацию возбуждений, усвоение и трансформацию ритма возбуждений, следовые процессы и легкую утомляемость.
Односторонность проведения возбуждения в нервных центрах связана с тем, что в синапсах нервные импульсы проходят только в одном направлении — от синаптиче-ского окончания аксона одного нейрона через синаптиче-скую щель на клеточное тело и дендриты других нейронов.
Замедление движения нервных импульсов связано с тем, что «телеграфный», т. е. электрический, способ передачи нервных импульсов в синапсах сменяется химическим, или медиаторным, скорость которого в тысячу раз меньше. Время этой так называемой синаптической задержки импульсов складывается из времени прихода импульса в синаптическое окончание, времени диффузии медиатора в синаптическую щель и его движения к постси-наптической мембране, времени изменения ионной проницаемости мембраны и возникновения потенциала действия, т. е. нервного импульса. В среднем все время передачи нервного импульса (потенциала действия) от одного нейрона через синапс к другому нейрону составляет около 1,5 мс.
В действительности в осуществлении какой-либо реакции человека участвуют сотни и тысячи нейронов и суммарное время задержки проведения нервных импульсов, называемое центральным временем проведения, увеличивается до сотен и более миллисекунд. Например, время реакции водителя с момента включения красного света светофора до начала его ответных действий будет составлять не менее 200 мс.
Таким образом, чем больше синапсов на пути движения нервных импульсов, тем больше проходит времени от начала раздражения до начала ответной реакции. Это время называют временем реакции или латентным временем рефлекса.
У детей время центральной задержки больше, оно увеличивается также при различных воздействиях на организм человека. При утомлении водителя оно может превышать 1000 мс, что приводит в опасных ситуациях к замедленным реакциям и дорожным авариям.
Суммация возбуждений была открыта И. М. Сеченовым в 1863 г. В настоящее время различают пространственную и временную суммацию нервных импульсов. Первая наблюдается при одновременном поступлении к одному нейрону нескольких импульсов, каждый из которых в отдельности является подпороговым раздражителем и не вызывает возбуждение нейрона. В сумме же нервные импульсы достигают необходимой силы и вызывают появление потенциала действия.
Временная суммация возникает при поступлении к постсинаптической мембране нейрона серии импульсов, в отдельности не вызывающих возбуждение нейрона. Сумма этих импульсов достигает пороговой величины раздражения и вызывает возникновение потенциала действия.
Явление суммации можно наблюдать, например, при одновременном подпороговом раздражении нескольких ре-цепторных зон кожи или при ритмическом подпороговом раздражении одних и тех же рецепторов. И в том и другом случае подпороговые раздражения вызовут ответную рефлекторную реакцию.
Усвоение и трансформация ритма возбуждений в нервных центрах были изучены известным русским и советским ученым А. А. Ухтомским (1875—1942) и его учениками. Сущность усвоения ритма возбуждений заключается в способности нейронов «настраиваться» на ритм поступающих раздражений, что имеет большое значение для оптимизации взаимодействия различных нервных центров при организации поведенческих актов человека. С другой стороны, нейроны способны трансформировать (изменять) поступающие к ним ритмические раздражения в свой собственный ритм.
После прекращения действия раздражителя активность нейронов, составляющих нервные центры, не прекращается. Время этого последействия, или следовых процессов, сильно варьирует у различных нейронов и в зависимости от характера раздражителей. Предполагают, что явление последействия имеет важное значение в понимании механизмов памяти. Непродолжительное последействие до 1 ч, вероятно, связано с механизмами краткосрочной памяти, а более длительные следы, хранящиеся в нейронах многие годы и имеющие большое значение в обучении детей и подростков, связаны с механизмами долговременной памяти.
Наконец, последняя особенность нервных центров — их быстрая утомляемость — также связана в значительной степени с «деятельностью синапсов. Существуют данные, что длительные раздражения приводят к постепенному истощению в синапсах запасов медиаторов, к снижению. чувствительности к ним постсинаптической мембраны. В результате рефлекторные ответные реакции начинают ослабевать и в конечном итоге полностью прекращаются.
НЕРВНЫЙ ИМПУЛЬС
Нервный импульс обеспечивает передачу информации от рецепторов к нервным центрам и от них к исполнительным органам — скелетной мускулатуре, гладким мышцам внутренних органов и сосудов, железам внутренней и внешней секреции и т. д.
Сложная информация о действующих на организм раздражениях кодируется в виде отдельных групп Нервных импульсов — рядов. Согласно закону «Все или ничего» (см.) амплитуда и длительность отдельных Нервных импульсов, проходящих по одному и тому же волокну, постоянны, а частота и количество Нервных импульсов в ряду зависят от интенсивности раздражения. Такой способ передачи информации является наиболее помехоустойчивым, т. е. в широких пределах не зависит от состояния проводящих волокон.
Распространение Нервных импульсов отождествляется с проведением потенциалов действия (см. Биоэлектрические потенциалы). Возникновение возбуждения может быть результатом раздражения (см.), напр, воздействие света на зрительный рецептор, звука на слуховой рецептор, или процессов, протекающих в тканях (спонтанное возникновение Н. и.). В этих случаях Н. и. обеспечивают согласованную работу органов при протекании какого-либо физиологического процесса (напр., в процессе дыхания Н. и. вызывают сокращение скелетных мышц и диафрагмы, результатом чего являются вдох и выдох, и т. д.).
В живых организмах передача информации может осуществляться и гуморальным путем, посредством выброса в русло крови гормонов, медиаторов и т. п. Однако преимущество информации, передаваемой при помощи Н. и., состоит в том, что она более целенаправленна, передается быстро и может быть точнее закодирована, чем сигналы, посылаемые гуморальной системой.
Факт, что нервные стволы являются путем, по к-рому передаются влияния от мозга к мышцам и в обратном направлении, был известен еще в эпоху античности. В средние века и вплоть до середины 17 в. считалось, что по нервам распространяется некая субстанция, подобная жидкости или пламени. Идея о электрической природе Н. и. возникла в 18 в. Первые исследования электрических явлений в живых тканях, связанных с возникновением и распространением возбуждения, были осуществлены Л. Гальвани. Г. Гельмгольц показал, что скорость распространения Н. и., к-рую ранее считали близкой к скорости света, имеет конечное значение и может быть точно измерена. Германн (L. Hermann) ввел в физиологию понятие потенциала действия. Объяснение механизма возникновения и проведения возбуждения стало возможным после создания С. Аррениусом теории электролитической диссоциации. В соответствии с этой теорией Бернштейн (J. Bernstein) предположил, что возникновение и проведение Н. и. обусловлено перемещением ионов между нервным волокном и окружающей средой. Англ. исследователи А. Ходжкин, Б. Катц и Э. Хаксли детально исследовали трансмембранные ионные токи, лежащие в основе развития потенциала действия. Позже стали интенсивно изучаться механизмы работы ионных каналов, по к-рым происходит обмен ионами между аксоном и окружающей средой, и механизмы, обеспечивающие способность нервных волокон проводить ряды Н. и. разного ритма и продолжительности.
Н. и. распространяется за счет местных токов, возникающих между возбужденным и невозбужденным участками нервного волокна. Ток, выходящий из волокна наружу в покоящемся участке, служит раздражителем. Наступающая после возбуждения в данном участке нервного волокна рефрактерность обусловливает поступательное движение Н. и.
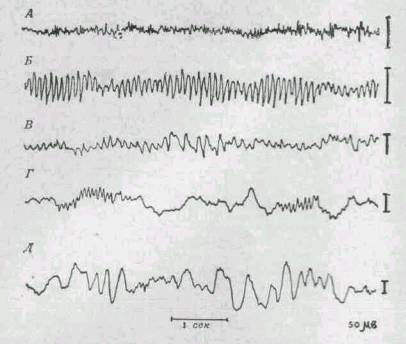
Количественно соотношения разных фаз развития потенциала действия можно охарактеризовать, сопоставляя их по амплитуде и длительности во времени. Так, напр., для миелиновых нервных волокон группы А млекопитающих диаметр волокна находится в пределах 1—22 мк, скорость проведения — 5—120 м/сек, длительность и амплитуда высоковольтной части (пика, или спайка) — 0,4—0,5 мсек и 100—120 мв соответственно, следовой негативный потенциал — 12—20 мсек (3—5% от амплитуды спайка), следовой позитивный потенциал — 40—60 мсек (0,2% от амплитуды спайка).
Возможности передачи разнообразной информации расширяются за счет повышения скорости развития потенциала действия, скорости распространения, а также за счет повышения лабильности (см.) — т. е. способности возбудимого образования воспроизводить в единицу времени высокие ритмы возбуждения.
Возникновение Н. и. в нервных клетках (см.) или рецепторах (см.) связано с деполяризацией мембраны, т. е. со снижением величины электрического потенциала на мембране (потенциала покоя, или мембранного потенциала). Если величина мембранного потенциала снижается на 10—20% (пороговый критический уровень), то местный процесс переходит в распространяющийся — возникает потенциал действия (см. Возбуждение).
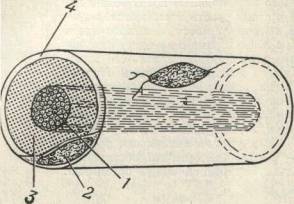
Конкретные особенности распространения Н. и. связаны со строением нервных волокон (см.). Сердцевина волокна (аксоплазма) обладает низким сопротивлением и, соответственно, хорошей проводимостью, а окружающая аксоплазму плазматическая мембрана — большим сопротивлением. Особенно велико электрическое сопротивление наружного слоя у миелинизированных волокон, у к-рых свободны от толстой миелиновой оболочки только перехваты Ранвье. В безмиелиновых волокнах Н. и. движется непрерывно, а в миелиновых — скачкообразно (сальтаторное проведение).
Различают декрементное и бездекрементное распространение волны возбуждения. Декрементное проведение, т. е. проведение возбуждения с угасанием, наблюдается в безмиелиновых волокнах. В таких волокнах скорость проведения Н. и. невелика и по мере отдаления от места раздражения раздражающее действие местных токов постепенно уменьшается вплоть до полного угасания. Декрементное проведение свойственно волокнам, иннервирующим внутренние органы, обладающие низкой функц, подвижностью. Без декрементное проведение характерно для миелиновых и тех безмиелиновых волокон, к-рые передают сигналы к органам, обладающим высокой реактивностью (напр., сердечной мышце). При бездекрементном проведении Н. и. проходит весь путь от места раздражения до места реализации информации без затухания.
Максимальная скорость проведения Н. и., зарегистрированная в быстропроводящих нервных волокнах млекопитающих, составляет 120 м/сек. Высокие скорости проведения импульса могут быть достигнуты за счет увеличения диаметра нервного волокна (у безмиелиновых волокон) или за счет повышения степени миелинизации. Распространение одиночного Н. и. само по себе не требует непосредственных энергетических затрат, т. к. при определенном уровне поляризации мембраны каждый участок нервного волокна находится в состоянии готовности к проведению и раздражающий стимул играет роль «спускового курка». Однако восстановление исходного состояния нервного волокна и поддержание его в готовности к проведению нового Н. и. связано с затратой энергии биохимических реакций, протекающих в нервном волокне. Процессы восстановления приобретают большое значение в случае проведения рядов Н. и. При проведении ритмического возбуждения (рядов импульсов) в нервных волокнах приблизительно вдвое возрастает теплопродукция и потребление кислорода, расходуются макроэргические фосфаты и повышается активность Na,K-АТФ-азы к-рую отождествляют с натриевым насосом. Изменение интенсивности протекания различных физ.-хим. и биохимических процессов зависит от характера ритмического возбуждения (продолжительность рядов импульсов и частота их следования) и физиологического состояния нерва. При проведении большого числа Н. и. в высоком ритме в нервных волокнах может накапливаться «метаболический долг» (это находит отражение в увеличении суммарных следовых потенциалов), и тогда процессы восстановления затягиваются. Но и в этих условиях способность нервных волокон проводить Н. и. долгое время остается неизменной.
Передача Н. и. с нервного волокна на мышечное или какой-либо другой эффектор осуществляется через синапсы (см.). У позвоночных животных в подавляющем большинстве случаев передача возбуждения на эффектор происходит при помощи выделения ацетилхолина (нервно-мышечные синапсы скелетной мускулатуры, синаптические соединения в сердце и др.). Для таких синапсов характерно строго одностороннее проведение импульса и наличие временной задержки передачи возбуждения.
В синапсах, в синаптической щели которых сопротивление электрическому току благодаря большой площади контактирующих поверхностей мало, происходит электрическая передача возбуждения. В них нет синаптической задержки проведения и возможно двустороннее проведение. Такие синапсы свойственны беспозвоночным животным.
Регистрация Н. и. нашла широкое применение в биол, исследованиях и клин, практике. Для регистрации используют шлейфные и чаще катодные осциллографы (см. Осциллография). При помощи микроэлектродной техники (см. Микроэлектродный метод исследования) регистрируют Н. и. в одиночных возбудимых образованиях — нейронах и аксонах. Возможности исследования механизма возникновения и распространения Н. и. значительно расширились после разработки метода фиксации потенциала. Этим методом были получены основные данные о ионных токах (см. Биоэлектрические потенциалы).
Нарушение проведения Н. и. происходит при повреждении нервных стволов, напр, при механических травмах, сдавливании в результате разрастания опухоли или при воспалительных процессах. Такие нарушения проведения Н. и. зачастую бывают необратимы. Следствием прекращения иннервации могут быть тяжелые функциональные и трофические расстройства (напр., атрофия скелетных мышц конечностей после прекращения поступления Н. и. вследствие необратимой травмы нервного ствола). Обратимое прекращение проведения Н. и. может быть вызвано специально, в терапевтических целях. Напр., с помощью анестезирующих средств блокируют импульсацию, идущую от болевых рецепторов в ц. н. с. Обратимое прекращение проведения Н. и. вызывает и новокаиновая блокада. Временное прекращение передачи Н. и. по нервным проводникам наблюдается и во время общего наркоза.
Библиография: Бpеже М. А. Электрическая активность нервной системы, пер. с англ., М., 1979; Жуков Е. К. Очерки по нервно-мышечной физиологии, Л., 1969; Коннели К. Восстановительные процессы и обмен веществ в нерве, в кн.: Совр, пробл. биофизики, пер. с англ., под ред. Г. М. Франка и А. Г. Пасынского, т. 2, с. 211, М., 1961; Костюк П. Г. Физиология центральной нервной системы, Киев, 1977; Латманизова Л. В. Очерк физиологии возбуждения, М., 1972; Общая физиология нервной системы, под ред. П. Г. Костюка, Л., 1979; Тасаки И. Нервное возбуждение, пер. с англ., М., 1971; Ходжкин А. Нервный импульс, пер. с англ., М., 1965; Ходоров Б. И. Общая физиология возбудимых мембран, М., 1975.
Распространение нервных импульсов
Нервные импульсы распространяются при перемещении ионов через мембрану нервной клетки и передаются из одной нервной клетки в другую с помощью нейромедиаторов.
В результате эволюции нервной системы человека и других животных возникли сложные информационные сети, процессы в которых основаны на химических реакциях. Важнейшим элементом нервной системы являются специализированные клетки нейроны. Нейроны состоят из компактного тела клетки, содержащего ядро и другие органеллы. От этого тела отходит несколько разветвленных отростков. Большинство таких отростков, называемых дендритами, служат точками контакта для приема сигналов от других нейронов. Один отросток, как правило самый длинный, называется аксоном и передает сигналы на другие нейроны. Конец аксона может многократно ветвиться, и каждая из этих более мелких ветвей способна соединиться со следующим нейроном.
Во внешнем слое аксона находится сложная структура, образованная множеством молекул, выступающих в роли каналов, по которым могут поступать ионы — как внутрь, так и наружу клетки. Один конец этих молекул, отклоняясь, присоединяется к атому-мишени. После этого энергия других частей клетки используется на то, чтобы вытолкнуть этот атом за пределы клетки, тогда как процесс, действующий в обратном направлении, вводит внутрь клетки другую молекулу. Наибольшее значение имеет молекулярный насос, который выводит из клетки ионы натрия и вводит в нее ионы калия (натрий-калиевый насос).
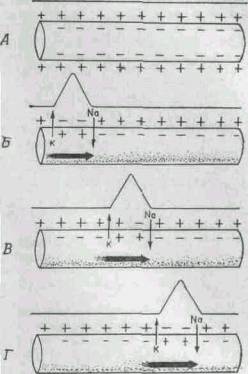
Когда клетка находится в покое и не проводит нервных импульсов, натрий-калиевый насос перемещает ионы калия внутрь клетки и выводит ионы натрия наружу (представьте себе клетку, содержащую пресную воду и окруженную соленой водой). Из-за такого дисбаланса разность потенциалов на мембране аксона достигает 70 милливольт (приблизительно 5% от напряжения обычной батарейки АА).
Однако при изменении состояния клетки и стимуляции аксона электрическим импульсом равновесие на мембране нарушается, и натрий-калиевый насос на короткое время начинает работать в обратном направлении. Положительно заряженные ионы натрия проникают внутрь аксона, а ионы калия откачиваются наружу. На мгновение внутренняя среда аксона приобретает положительный заряд. При этом каналы натрий-калиевого насоса деформируются, блокируя дальнейший приток натрия, а ионы калия продолжают выходить наружу, и исходная разность потенциалов восстанавливается. Тем временем ионы натрия распространяются внутри аксона, изменяя мембрану в нижней части аксона. При этом состояние расположенных ниже насосов меняется, способствуя дальнейшему распространению импульса. Резкое изменение напряжения, вызванное стремительными перемещения ионов натрия и калия, называют потенциалом действия. При прохождении потенциала действия через определенную точку аксона, насосы включаются и восстанавливают состояние покоя.
Потенциал действия распространяется довольно медленно — не более доли дюйма за секунду. Для того чтобы увеличить скорость передачи импульса (поскольку, в конце концов, не годится, чтобы сигнал, посланный мозгом, достигал руки лишь через минуту), аксоны окружены оболочкой из миелина, препятствующей притоку и оттоку калия и натрия. Миелиновая оболочка не непрерывна — через определенные интервалы в ней есть разрывы, и нервный импульс перескакивает из одного «окна» в другое, за счет этого скорость передачи импульса возрастает.
Когда импульс достигает конца основной части тела аксона, его необходимо передать либо следующему нижележащему нейрону, либо, если речь идет о нейронах головного мозга, по многочисленным ответвлениям многим другим нейронам. Для такой передачи используется абсолютно иной процесс, нежели для передачи импульса вдоль аксона. Каждый нейрон отделен от своего соседа небольшой щелью, называемой синапсом. Потенциал действия не может перескочить через эту щель, поэтому нужно найти какой-то другой способ для передачи импульса следующему нейрону. В конце каждого отростка имеются крошечные мешочки, называющие (пресинаптическими) пузырьками, в каждом из которых находятся особые соединения — нейромедиаторы. При поступлении потенциала действия из этих пузырьков высвобождаются молекулы нейромедиаторов, пересекающие синапс и присоединяющиеся к специфичным молекулярным рецепторам на мембране нижележащих нейронов. При присоединении нейромедиатора равновесие на мембране нейрона нарушается. Сейчас мы рассмотрим, возникает ли при таком нарушении равновесия новый потенциал действия (нейрофизиологи продолжают искать ответ на этот важный вопрос до сих пор).
После того как нейромедиаторы передадут нервный импульс от одного нейрона на следующий, они могут просто диффундировать, или подвергнуться химическому расщеплению, или вернуться обратно в свои пузырьки (этот процесс нескладно называется обратным захватом). В конце XX века было сделано поразительное научное открытие — оказывается, лекарства, влияющие на выброс и обратный захват нейромедиаторов, могут коренным образом изменять психическое состояние человека. Прозак (Prozac*) и сходные с ним антидепрессанты блокируют обратный захват нейромедиатора серотонина. Складывается впечатление, что болезнь Паркинсона взаимосвязана с дефицитом нейромедиатора допамина в головном мозге. Исследователи, изучающие пограничные состояния в психиатрии, пытаются понять, как эти соединения влияют на человеческий рассудок.
По-прежнему нет ответа на фундаментальный вопрос о том, что же заставляет нейрон инициировать потенциал действия — выражаясь профессиональным языком нейрофизиологов, неясен механизм «запуска» нейрона. В этом отношении особенно интересны нейроны головного мозга, которые могут принимать нейромедиаторы, посланные тысячей соседей. Об обработке и интеграции этих импульсов почти ничего не известно, хотя над этой проблемой работают многие исследовательские группы. Нам известно лишь, что в нейроне осуществляется процесс интеграции поступающих импульсов и выносится решение, следует или нет инициировать потенциал действия и передавать импульс дальше. Этот фундаментальный процесс управляет функционированием всего головного мозга. Неудивительно, что эта величайшая загадка природы остается, по крайней мере сегодня, загадкой и для науки!
К. Вили Нервная система
 |
 | |||||
 | ПЕРЕДАЧА В СИНАПСЕ
|